Curso interactivo de Sistemática Filogenética
![]()
![]()
![]()
Tema 4.- La construcción de cladogramas. Ejemplos prácticos.
Vamos a abordar ahora casos prácticos de construcción de cladogramas, utilizando ejemplos reales. Comenzaremos por el ejemplo que utilizamos en el primer tema y que se ha convertido en un clásico a la hora de comparar las clasificaciones evolutivas y filogenéticas: las relaciones de reptiles escamosos (lagartos y serpientes), cocodrilos y aves.
Algunas de las semejanzas entre cocodrilos y escamosos (y diferencias con las aves) se recogen en esta tabla:
| Reptiles escamosos | Cocodrilos | Aves |
| Escamas córneas | Escamas córneas | Plumas |
| Patas anteriores marchadoras | Patas anteriores marchadoras | Alas |
| Mandíbulas normales | Mandíbulas normales | Mandíbulas formando un pico |
| Ectotermos | Ectotermos | Endotermos ("sangre caliente") |
Se podrían citar otras semejanzas, pero basta con estas. Un primer análisis mostrará que todas ellas, sin excepción, son simplesiomorfías (recordemos, caracteres derivados de un ancestral lejano y común a todo el grupo). El análisis de grupos externos (anfibios, tortugas...) permite encontrar sin dificultad las escamas córneas, las patas anteriores marchadoras, dientes y ectotermia. Luego, de acuerdo con lo que hemos visto acerca de las simplesiomorfías, todas estas semejanzas heredadas de ancestrales remotos no indican relación de parentesco.
En cambio sí que es posible encontrar caracteres derivados (apomorfos) y compartidos por cocodrilos y aves, es decir, sinapomorfías. En concreto:
| Dientes tecodontos (en alvéolos)* |
| Ventana preorbitaria** |
| Pelvis modificada*** |
| Tabicación completa del ventrículo cardiaco y reducción o desaparición del arco aórtico izquierdo**** |
*Presentes en las aves primitivas (Archaeopteryx).
Desaparecen en las aves modernas.
**Presente en los antepasados de los cocodrilos. Desaparece en cocodrilos
modernos.
***Consiste en la curvatura hacia abajo de pubis e isquion. A veces el pubis
se dirige hacia atrás o desaparece.
****Existe un pequeńo orificio (foramen de Panizza) en cocodrilos.
No existe ningún carácter derivado y compartido por aves y reptiles escamosos. Por ello, el cladograma que mejor refleja este análisis de caracteres es el siguiente:
 En realidad sabemos que este cladograma, que
hemos obtenido por medio del análisis de caracteres, se ajusta perfectamente a lo que se
conoce acerca de la filogenia de los reptiles y del origen de las aves. En efecto, tanto
las aves como los cocodrilos son los únicos supervivientes de determinados grupos
de dinosaurios, mientras que los reptiles escamosos, aunque emparentados con dinosaurios,
cocodrilos y aves, comparten ancestrales comunes más alejados en el tiempo.
En realidad sabemos que este cladograma, que
hemos obtenido por medio del análisis de caracteres, se ajusta perfectamente a lo que se
conoce acerca de la filogenia de los reptiles y del origen de las aves. En efecto, tanto
las aves como los cocodrilos son los únicos supervivientes de determinados grupos
de dinosaurios, mientras que los reptiles escamosos, aunque emparentados con dinosaurios,
cocodrilos y aves, comparten ancestrales comunes más alejados en el tiempo.
żQue consencuencias tiene para la clasificación de los vertebrados esta forma de realizar el análisis filogenético? La más evidente es que, dado que sólo podemos establecer taxones monofiléticos, no resulta aceptable un taxón Reptiles. Ha quedado claro que los reptiles no constituyen un grupo monofilético, definido por sinapomorfías. En efecto, no hay sinapomorfías que unan a cocodrilos y lagartos, como antes hemos expuesto, pero tampoco las hay entre tortugas y lagartos, por ejemplo. Sin embargo, cocodrilos, dinosaurios y aves sí pueden ser reunidos en un taxón definido por las sinapomorfías antes citadas y por otras. Dicho taxón se denomina Arcosaurios.
Vamos a complicar un poco la cuestión, intentando insertar, en el cladograma anterior, a quelonios y mamíferos, para tener de esta forma representantes de los principales grupos actuales de amniotas. Para ello vamos a manejar la siguiente tabla de caracteres, utilizando a los anfibios como grupo externo:
| Quelonios | Mamíferos | Escamosos | Cocodrilos | Aves | Anfibios (grupo externo) | |
| Aberturas temporales en el cráneo | No tienen (anápsidos) |
Un par (sinápsidos) |
Dos pares (diápsidos) |
Dos pares (diápsidos) |
Dos pares (diápsidos) |
No tienen |
| Ventana preorbitaria | Ausente | Ausente | Ausente | Presente* | Presente | Ausente |
| Regulación de la temperatura | Ectotermos | Endotermos | Ectotermos | Ectotermos | Endotermos | Ectotermos |
| Pelvis | Normal | Normal | Normal | Modificada | Modificada | Normal |
| Arco aórtico principal | Izq. y Derecho | Izquierdo | Izq. y Derecho | Derecho | Derecho | Izq. y Derecho |
*En los antepasados de los cocodrilos, como ya hemos dicho antes.
- Un par de aberturas temporales en el cráneo*.
- Dos pares de aberturas temporales en el cráneo*.
- Presencia de ventana preorbitaria
- Endotermia.
- Pelvis modificada.
- Sólo arco aórtico izquierdo*.
- Sólo arco aórtico derecho*.
*A partir del criterio del grupo externo, no podemos decidir si uno de estos caracteres es derivado respecto del otro, o si ambos son independientes. Lo que está claro es que ambos son derivados respecto de la ausencia de ventanas o del doble arco aórtico.
Son derivados y no compartidos (autopomorfías) la presencia de un par de aberturas temporales y la posesión de arco aórtico izquierdo (condiciones ambas de mamíferos). Por tanto no nos servirán para establecer relaciones de parentesco. Si utilizamos como sinapomorfías los restantes, podremos generar no uno, sino muchos cladogramas, de los que trabajaremos con estos tres:
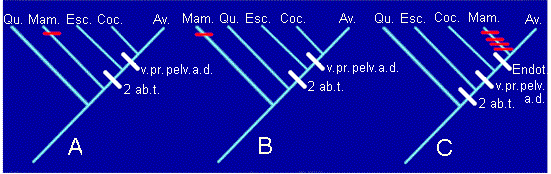
2 ab.t.= 2 aberturas temporales. v.pr.=ventana preorbitaria. pelv.=pelvis
modificada. a.d.=arco aórtico derecho. Endot=endotermia.
Podemos comprobar como en los cladogramas A y B asumimos que la endotermia apareció de forma independiente en Mamíferos y Aves (convergencia). En cambio, el cladograma C asume que la endotermia estaba presente en el ancestral común de Mamíferos y Aves, pero en este caso hay que aceptar que en el linaje de los mamíferos el número de aberturas temporales pasa de dos a una, se pierde la pelvis modificada, la ventana preorbitaria y se sustituye el arco aórtico derecho por el izquierdo (reversiones). Cada uno de estos cambios (convergencias o reversiones) se representa por una raya roja en el cladograma.
żPor cuál optaremos? En los casos en los que la distribución de caracteres en los taxones pueda explicarse por más de un cladograma (en realidad esto incluye a todos los casos posibles), aplicaremos el llamado criterio de máxima parsimonia. Este criterio aconseja a elegir aquel cladograma más sencillo, más simple, es decir, el que implique el menor grado de homoplasia. La homoplasia se puede definir como la existencia de similitud no debida a parentesco. El grado de homoplasia es el número de convergencias o reversiones que es preciso asumir a la hora de construir el cladograma. En el caso anterior, los cladogramas A y B tienen una convergencia (no dos, ˇojo!, la convergencia se produce cuando la endotermia aparece por segunda vez). El cladograma C tiene cuatro reversiones (número de aberturas temporales, pelvis, ventana preorbitaria y arco aórtico derecho).
En conclusión, el criterio de máxima parsimonia nos haría descartar el cladograma C. Sin embargo, no podemos decidir entre el A y el B, puesto que ambos están igualmente justificado por sinapomorfías y por grado de homoplasia. Esta situación es frecuente en Sistemática Filogenética. Es necesario buscar más sinapomorfías que nos permitan decidir entre las dos opciones A y B. De hecho, si hubiéramos utilizado más caracteres hubiéramos terminado decantándonos por el cladograma B.
Vamos a utilizar el ejemplo anterior para ilustrar algunas cuestiones importantes de la Sistemática Filogenética. Por ejemplo la equivalencia potencial de convergencias y reversiones que justifica el término común de homoplasia. Observemos el siguiente cladograma:
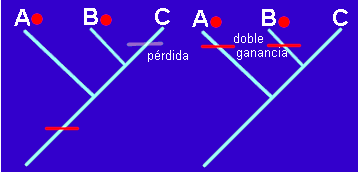 A y B comparten un carácter apomorfo cualquiera simbolizado
por el punto rojo. Sin embargo A y B no están más emparentados que B y C, que son grupos
hermanos por compartir más caracteres derivados (no representados en la figura). El
carácter "punto rojo" es, por tanto, un caso de homoplasia. Sin embargo, no es
posible decidir entre los dos escenarios mostrados en la figura, una reversión (a la
izquierda) o una convergencia (a la derecha). Cualquiera de las dos posibilidades explica
la homoplasia de forma igualmente parsimoniosa.
A y B comparten un carácter apomorfo cualquiera simbolizado
por el punto rojo. Sin embargo A y B no están más emparentados que B y C, que son grupos
hermanos por compartir más caracteres derivados (no representados en la figura). El
carácter "punto rojo" es, por tanto, un caso de homoplasia. Sin embargo, no es
posible decidir entre los dos escenarios mostrados en la figura, una reversión (a la
izquierda) o una convergencia (a la derecha). Cualquiera de las dos posibilidades explica
la homoplasia de forma igualmente parsimoniosa.
Si hemos entendido el concepto de homoplasia, entenderemos también el concepto de "longitud del árbol" (treelength en inglés). Imaginemos cada uno de los cambios como un "paso" que ańade una unidad de longitud al árbol. En el caso de nuestro cladograma anterior (elimino a los quelonios para simplificar) tenemos:
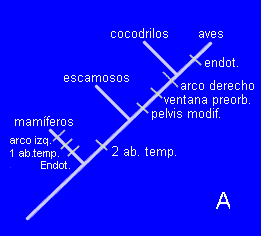
El equivalente al árbol A requiere un total de 8 cambios para explicar la distribución de los caracteres. Por tanto, la longitud del árbol será de 8 pasos. Comprueba que la hipótesis B implica un árbol de la misma longitud.
(observa que la endotermia aparece dos veces)
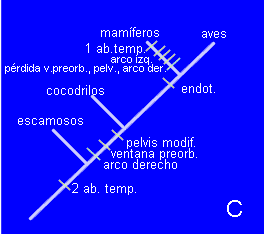
En cambio, el equivalente del árbol C requiere un mínimo de 10 cambios para explicar la distribución de caracteres en los taxones. Su longitud será de 10 pasos.
(la posición de los mamíferos explica la presencia de endotermia, pero a cambio es preciso asumir la pérdida de la ventana preorbitaria, la pelvis modificada, el arco aórtico derecho y un par de aberturas temporales).
La longitud del árbol nos proporciona otra forma de enunciar el criterio de parsimonia. Elegiremos aquel cladograma que implique una menor longitud, es decir, que implique el menor número de cambios. Por otro lado podemos expresar esto de forma cuantitativa. En efecto, podemos definir un índice que relacione el número mínimo de cambios posible con el número real de cambios. Esto es lo que se denomina el Indice de consistencia. En el ejemplo anterior, el número mínimo de cambios en los caracteres es de siete, los mismos que se detallan en la tabla de arriba (si no los recuerdas pulsa aquí).
Así, los árboles correspondientes a las hipótesis A y B tiene una longitud de 8 pasos sobre un mínimo de 7. El Indice de consistencia será 7/8=0,875.
El árbol correspondiente a la hipótesis C tiene una longitud de 10 pasos sobre el mínimo de 7. Su Indice de consistencia será 7/10=0,7.
Aquí tenemos otra formulación del criterio de parsimonia: Entre dos cladogramas posibles elegiremos el que presente un mayor Indice de consistencia. Podemos comprobar cómo no pudimos en su momento decidir entre las hipótesis A y B, porque ambas presentaban el mismo grado de consistencia. En cambio, la consistencia de la opción C, y de cualquier otro cladograma alternativo, siempre será inferior a 0.875.
ˇBien, y ha llegado el momento de pasar a los ejercicios!
