Curso interactivo de Sistemática Filogenética
![]()
![]()
![]()
Tema 5.- Cladogramas y clasificación.
En este tema veremos cómo es posible aplicar el método filogenético al sistema linneano, un sistema que fue diseńado en un contexto preevolutivo, un contexto muy diferente al que estamos describiendo aquí.
Antes de desarrollar el tema, es preciso comprobar que habéis comprendido una propiedad importante de la Sistemática Filogenética. Un mismo carácter puede ser sinapomorfía, simplesiomorfía y autopomorfía a diferentes niveles taxonómicos. żCómo es esto posible? Vamos a verlo con un ejemplo.
Tomemos como ejemplo el carácter "presencia de vértebras". Este carácter no es apomorfo ni plesiomorfo en sí mismo, sino en relación a un contexto taxonómico determinado. Veamos tres situaciones:
- Estamos trabajando sobre las relaciones filogenéticas de los Deuteróstomos, es decir, Equinodermos, Hemicordados, Urocordados, Cefalocordados y Vertebrados. Es obvio que la presencia de vértebras será un carácter derivado y no compartido (sólo tienen vértebras los vertebrados). Por tanto será una autopomorfía.
- Trabajamos ahora con las relaciones filogenéticas de los siguientes taxones: Urocordados, Cefalocordados, Elasmobranquios, Actinopterigios, Tetrápodos. La presencia de vértebras se da en tres de estos taxones (Elasmobranquios, Actinopterigios, Tetrápodos). Por tanto es un carácter derivado y compartido (una sinapomorfía).
- Ahora estamos a otro nivel, examinando las relaciones filogenéticas entre Lisanfibios, Lepidosaurios, Cocodrilos, Aves y Mamíferos, con los Elasmobranquios como grupo externo. Las vértebras han pasado a ser un carácter primitivo y compartido por todos los taxones, es decir, una simplesiomorfía.
Las tres posibilidades se reflejan en los siguientes cladogramas:
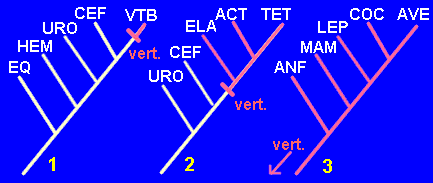
Como puede comprobarse, sólo en el caso 2, en el que la presencia de vértebras se convierte en una sinapomorfía, resulta informativa, es decir, útil para la construcción del cladograma.
Una vez que esto queda claro, podemos entrar en la cuestión de cómo debemos clasificar. La primera conclusión, que ya exponíamos en el tema 2, es que sólo los grupos estrictamente monofiléticos pueden ser considerados como taxones. Entonces definimos a los grupos monofiléticos como "todos y cada uno de los descendientes de un ancestral común". Ahora podemos ańadir que el grupo monofilético es el definido por sinapomorfías, puesto que son las sinapomorfías las que nos hacen elaborar una hipótesis sobre un ancestral común a todos los miembros de un taxón. De esta forma hacemos que la definición descanse sobre nuestra herramienta de trabajo, la sinapomorfía.
żSignifica esto que los taxones, al igual que los cladogramas y las sinapomorfías, son provisionales? Evidentemente, esto es así. Un taxón, en Sistemática Filogenética, ímplica una hipotesis de relaciones filogenéticas y las hipótesis son, por definición, provisionales. Por esto, el panorama que nos dibuja la Sistemática Filogenética es el de una clasificación dinámica, sujeta a cambios pero, al mismo tiempo, organizada de acuerdo con criterios objetivos. Cualquier propuesta de clasificación debe justificarse, tanto con sinapomorfías como en lo que se refiere al criterio de máxima parsimonia.
| Peces |
| Anfibios |
| Reptiles |
| Aves |
| Mamíferos |
Todo esto ha cambiado radicalmente algunos esquemas tradicionales de clasificación. Recordemos la clasificación clásica de los vertebrados, de raíz linneana, que se muestra en esta tabla. Desde el punto de vista de la Sistemática Filogenética sólo las Aves y los Mamíferos serían taxones aceptables, en tanto en cuanto son monofiléticos. Una clasificación coherente con la Sistemática Filogenética requiere el establecimiento de taxones estrictamente monofiléticos, como ya veremos. Lo mismo sucede con ciertas agrupaciones a nivel suprafilético (por encima del phylum), tales como "acelomados", "celomados" o "pseudocelomados", agrupaciones difícilmente justificables con sinapomorfías.
Antes de exponer un ejemplo de clasificación filogenética, es preciso discutir un punto que hemos pasado por alto. La Sistemática Filogenética proporciona un método de agrupación, pero en la clasificación no sólo consiste en agrupar sino también en asignar al grupo una determinada categoría taxonómica (familia, orden, subclase, etc.).
Es preciso reconocer que la Sistemática Filogenética no proporciona un criterio universal de asignación de categorías. La respuesta consiste en que lo importante es que la clasificación 1) se base en grupos monofiléticos, 2) refleje el parentesco filogenético en la medida de lo posible. Asignar a un grupo la categoría de superorden o subclase se convierte así en mucho menos relevante que establecer las relaciones filogenéticas de dicho grupo con grupos cercanos. No se nos escapa que dicha asignación sigue conteniendo un componente subjetivo muy grande. Podemos prever que el cladograma, por tanto, irá ganando importancia en los libros de texto y artículos sobre Sistemática, en detrimento de los listados "lineales", cuyo contenido informativo es mucho menor.
Veamos qué taxones podemos distinguir en los vertebrados vivientes de acuerdo con los criterios de la Sistemática Filogenética. Un cladograma posible (provisional y sujeto a revisión, como todos los cladogramas) sería el siguiente:
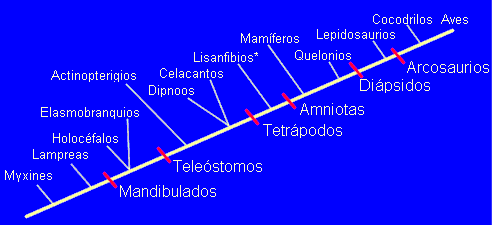
*Lisanfibios: anfibios actuales. Existen muchas incertidumbres en lo que se
refiere a la posición de holocéfalos, dipnoos, celacantos y quelonios.
Sólo se han indicado los principales taxones, mejor sustentados por
sinapomorfías. Dichas sinapomorfías son las siguientes:
| Taxón: | Sinapomorfías: |
| Mandibulados | Presencia de mandíbulas |
| Teleóstomos | Mandíbulas secundarias (formadas por huesos dérmicos, no branquiales) |
| Tetrápodos | Miembro quiridio. Cintura pélvica fusionada a la columna vertebral |
| Amniotas | Amnios y alantoides |
| Diápsidos | Dos pares de aberturas temporales en el cráneo |
| Arcosaurios | Pelvis modificada. Ventana preorbitaria |
Los clásicos taxones (peces, anfibios, reptiles) quedan así sustituidos por los nuevos. Es sin duda una revolución, pero żacaso no fue una revolución la obra de Linneo?
Todo lo que hemos dicho hasta ahora tiene una importante consecuencia cuando examinamos el problema de la especie. En la Sistemática Evolutiva, el concepto de especie era muy importante. Ya expusimos en el tema 1 que la Sistemática Evolutiva propugnaba la utilización del concepto biológico de especie en lugar del concepto tipológico típico del sistema linneano.
La importancia de establecer un criterio a la hora de definir la especie se debe a los procedimientos usados tanto en el esencialismo linneano como en Sistemática Evolutiva. En ambos casos se trata de utilizar a la especie como entidad objetiva (realmente existente en la naturaleza) y unidad de clasificación. Una vez delimitada la especie, el procedimiento consiste en agrupar especies en taxones de categoría cada vez mayor (Géneros, familias, órdenes, etc.).
En cambio, dado que la Sistemática Filogenética parte del establecimiento de hipótesis sobre las relaciones filogenéticas, y no de la delimitación de especies, parece claro que el debate sobre el concepto de especie no es fundamental en Sistemática Filogenética, o al menos no es tan fundamental como en otras escuelas taxonómicas. Un ejemplo nos ayudará a comprender este punto.
Recordemos que la Sistemática Filogenética nos ayuda a establecer hipótesis de parentesco filogenético a cualquier nivel taxonómico, supra o infraespecífico. Supongamos que hemos estudiado la filogenia de cuatro poblaciones de animales (1,2,3 y 4) muy emparentadas entre sí, poblaciones que en términos tradicionales podrían ser consideradas especies próximas o "subespecies". Fruto de nuestro estudio podría ser el siguiente cladograma:
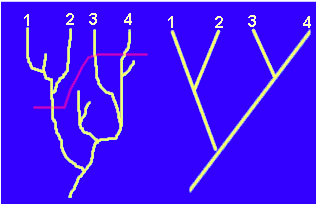 La
figura de la izquierda representa el proceso real de filogenia de las cuatro poblaciones.
Hemos seńalado en rojo el momento en que la progresiva divergencia de los linajes origina
la pérdida de la interfecundidad, es decir, individuos de las poblaciones 1 y 2 son
interfecundos, pero los individuos de las poblaciones 3 y 4 sólo pueden reproducirse con
los de la misma población.
La
figura de la izquierda representa el proceso real de filogenia de las cuatro poblaciones.
Hemos seńalado en rojo el momento en que la progresiva divergencia de los linajes origina
la pérdida de la interfecundidad, es decir, individuos de las poblaciones 1 y 2 son
interfecundos, pero los individuos de las poblaciones 3 y 4 sólo pueden reproducirse con
los de la misma población.
Si adoptamos el criterio de especie biológica, parece claro que estamos ante tres especies (1+2, 3 y 4). Si dicho criterio no fuera aplicable por cualquier razón, es posible que una cierta diferenciación morfológica pudiera llevar a interpretar las poblaciones 1 y 2 como dos especies diferentes. Queda claro, por tanto, que la elección del criterio de especie puede suponer cambios en la clasificación. Sin embargo, żla elección de un criterio de especie u otro supondría cambios en el cladograma de la derecha? Parece obvio que no.
No quiere esto decir que la delimitación de especies no sea importante, ni mucho menos. Para el biólogo es imprescindible saber a qué especie pertenece el organismo con el que está trabajando. Lo que queremos decir es que dicha tarea de delimitación específica se ha convertido en una empresa relativamente independiente de la determinación de relaciones de parentesco, relaciones que son las que finalmente van a nutrir la clasificación de los seres vivos.
