Curso interactivo de Sistemática Filogenética
![]()
![]()
![]()
Tema 3.- Los métodos de la Sistemática Filogenética. La inferencia filogenética. Caracteres apomorfos y plesiomorfos. Determinación de la polaridad de los caracteres.
En los temas anteriores hemos dejado claro que la Sistemática Filogenética utiliza las relaciones de parentesco filogenético como criterio a la hora de definir los taxones y de agruparlos en taxones de categoría superior. Esto quiere decir que el proceso fundamental consiste en hacer una inferencia filogenética, es decir, elaborar una hipótesis acerca del orden de aparición de los ancestrales comunes más recientes. Veamos un ejemplo de esto.
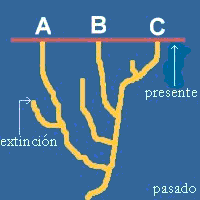 Supongamos tres taxones
cualesquiera A, B y C. Para poder trabajar, debemos suponer de entrada que son
estrictamente monofiléticos, es decir, que todos y cada uno de sus componentes derivan de
un ancestral común no compartido con otros taxones. Estos taxones son resultado de una
historia evolutiva que no podemos conocer con detalle. Pero supongamos por un momento que
sí podemos conocer cuál ha sido la historia de los linajes de A, B, y C, y que dicha
historia se representa en la figura adyacente.
Supongamos tres taxones
cualesquiera A, B y C. Para poder trabajar, debemos suponer de entrada que son
estrictamente monofiléticos, es decir, que todos y cada uno de sus componentes derivan de
un ancestral común no compartido con otros taxones. Estos taxones son resultado de una
historia evolutiva que no podemos conocer con detalle. Pero supongamos por un momento que
sí podemos conocer cuál ha sido la historia de los linajes de A, B, y C, y que dicha
historia se representa en la figura adyacente.
Volvamos a nuestro problema. Para tres taxones cualesquiera (A, B y C) pueden formularse tres y sólo hipótesis de parentesco que podemos representar de la siguiente forma (estos diagramas se denominan cladogramas, veremos luego qué son y cómo se construyen):
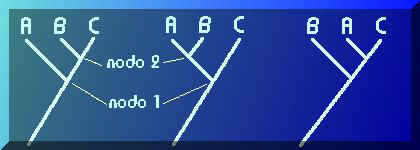
En el primer caso se propone que B y C están más emparentados porque su ancestral común (representado por el nodo 2 o unión de las dos ramas que llevan a B y C) es más reciente que el ancestral que ambos comparten con A (representado por el nodo 1). En el segundo caso los más emparentados son A y B, y en el tercer caso A y C. No hay más posibilidades.Observa que el orden de los nodos (que representan ancestrales comunes más recientes) nos indica el grado relativo de parentesco.
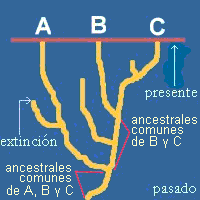
Si volvemos a la primera figura de este tema, la que representa lo que realmente ocurrió, nos decantaremos por la primera de las tres hipótesis. En efecto, B y C tienen toda una serie de ancestrales comunes que son más recientes que los ancestrales comunes que comparten A y B, por un lado, y A y C por el otro. Pero como no es posible conocer de forma cierta la filogenia (ˇa menos que tengamos el don de la clarividencia!), debemos buscar criterios objetivos que nos permitan elegir una de las tres hipótesis.
La clave nos la va a proporcionar el análisis de los caracteres de los organismos. Consideremos un organismo cualquiera, por ejemplo, un ratón. Podemos reconocer en el ratón caracteres que han tenido un origen más o menos remoto en el tiempo. Por ejemplo, sus incisivos de crecimiento continuo, típicos de los roedores, constituyen un carácter relativamente reciente. La presencia de pelos o glándulas mamarias, características de mamíferos, deben de tener un origen mucho más antiguo que el desarrollo de dichos incisivos especiales. Aún más antigua es la presencia de cráneo o vértebras, características de los vertebrados. Y todavía más remoto en el tiempo es la presencia de uniones intercelulares especializadas o colágeno (caracteres de metazoos) o núcleo celular (eucariotas). En Sistemática Filogenética se habla de caracteres apomorfos (recientes) o plesiomorfos (antiguos)
Caracteres son los atributos homólogos que utilizamos para establecer relaciones de parentesco (por ejemplo, número de dedos). Estados de carácter son las distintas variantes que pueden adoptar los caracteres (por ejemplo 1,2 ó 5 dedos). En la práctica, para facilitar las cosas, no haremos esta distinción en general y hablaremos de caracteres salvo que se indique lo contrario. |
De lo antes dicho se desprende que existe un orden temporal en la aparición de los caracteres o, utilizando una terminología más rigurosa, de los estados de los caracteres (ver recuadro). Como veremos a continuación, si podemos establecer un orden de aparición de determinados caracteres, si podemos distinguir entre estados ancestrales o plesiomorfos y estados avanzados o apomorfos, tendremos una herramienta para realizar inferencias filogenéticas.
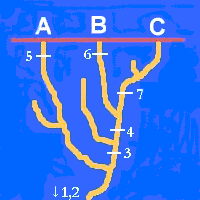
Volvamos a nuestro problema con los taxones A, B y C. En esta figura, la que representa lo que realmente sucedió, hemos expuesto la historia de algunos caracteres de los taxones. Podemos comprobar cómo los caracteres 1 y 2 se originaron en una etapa muy antigua, muy anterior a la diversificación de los linajes de A, B y C. Los caracteres 3 y 4, en cambio, aparecieron en el linaje del ancestral común de B y C. Por último los caracteres 5, 6 y 7 han aparecido en los linajes de los taxones A, B y C, respectivamente.
Si hemos entendido esto comprenderemos que el taxón A presente los caracteres 1,2
y 5. El taxón B posee los caracteres 1,2,3,4 y 6. Por último, los caracteres 1,2,3,4 y 7
estarán presentes en el taxón C.
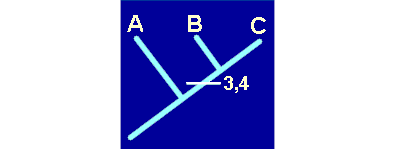
De acuerdo con esto, el hecho de que B y C compartan dos caracteres recientes, derivados o apomorfos (3 y 4), es una consecuencia directa de su parentesco, dado que recibieron dichos caracteres de un ancestral común, no compartido con A. En este caso hablaremos de una sinapomorfía. En cambio, que A, B y C compartan caracteres primitivos o plesiomorfos como el 1 ó el 2, heredados de un lejano ancestral común de todo el grupo, no ańade ninguna información. Esta compartición se denomina simplesiomorfía. Por último, tampoco nos informa de nada la posesión de caracteres derivados y no compartidos, como los 5, 6 y 7. A estos caracteres se les denomina autopomorfías.
Debe quedar claro, por tanto, que las sinapomorfías, los caracteres compartidos por dos o más taxones y derivados (aparecidos) dentro del grupo que estamos estudiando, son las que revelan relaciones significativas de parentesco.
Si hemos entendido esto, el problema se va a reducir, en la práctica a:
- Determinar las sinapomorfías, qué caracteres son derivados dentro del grupo y compartidos por dos o más taxones.
- Construir un diagrama que refleje la hipótesis de relaciones de parentesco a partir de dichas sinapomorfías.
Estos diagramas de relaciones de parentesco, que ya hemos presentado arriba, se llaman cladogramas. Cada uno de los nodos de un cladograma representa un hipotético ancestral común para los dos taxones que parten de dicho nodo (dichos taxones que nacen de un mismo nodo son grupos-hermanos. En los cladogramas deben siempre especificarse las sinapomorfías que justifican todos y cada uno de los nodos.
Bien, pues sólo nos queda, antes de pasar a la práctica, saber qué criterios podemos utilizar para determinar la polaridad de los caracteres, es decir, el sentido del cambio, cuál es el estado ancestral y cuál el derivado.
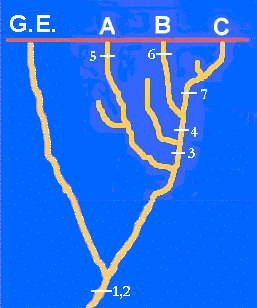
El criterio más utilizado es el de la comparación con grupos externos (out-groups). Volvamos a la figura del principio, ampliada para incluir otro linaje próximo. Podemos ver cómo los caracteres plesiomorfos, más antiguos (1,2) aparecen en grupos relacionados con el nuestro, pero cercanos a él (G.E. en la figura). Es evidente que los estados de carácter aparecidos dentro de nuestro grupo (3,4,5,6,7) no van a ser encontrados fuera de él, en un grupo externo. ˇEl carácter se "desplaza" hacia delante en el tiempo, no hacia atrás! Por tanto, el criterio nos dice que los estados plesiomorfos de carácter van a ser frecuentemente encontrados en grupos externos, mientras que los apomorfos no lo serán.
Un segundo criterio nos lo proporciona el desarrollo embrionario. En efecto, como notó el embriólogo K.E. Von Baer, los caracteres generales (plesiomorfos) aparecen siempre antes que los especiales (apomorfos). Por tanto la ontogenia de los caracteres refleja la polaridad del cambio.
Ya podemos resolver nuestro problema. Si examinamos la distribución de caracteres en los tres taxones A, B y C resolvemos:
- Descartar los caracteres 1 y 2 por ser simplesiomorfías.
- Descartar los caracteres 5, 6 y 7 por ser autopomorfías (derivados, pero no compartidos)
- Aceptar como sinapomorfías los caracteres 3 y 4, que asumimos como derivados dentro de nuestro grupo y compartidos por los taxones B y C.
- Por consiguiente, elegimos el primero de los tres cladogramas, ya que las sinapomorfías 3 y 4 revelan una posible relación de parentesco, un ancestral común a través del cual los taxones B y C recibieron los caracteres 3 y 4.
Vamos a hacer algunos ejercicios antes de entrar en mayores complicaciones.
