Curso interactivo de Sistemática Filogenética
![]()
![]()
Tema 1.- Introducción a la Sistemática. La Sistemática preevolutiva. La Nueva Sistemática. El Feneticismo.
 La
práctica de la Sistemática consiste en la nominación
y ordenamiento de los seres vivos de acuerdo con unas normas aceptadas
por toda la comunidad científica. En la actualidad, estas normas
constituyen un sistema denominado linneano en honor de Karl Von Linneo
(1707-1778). Dicho sistema se basa en dos principios:
La
práctica de la Sistemática consiste en la nominación
y ordenamiento de los seres vivos de acuerdo con unas normas aceptadas
por toda la comunidad científica. En la actualidad, estas normas
constituyen un sistema denominado linneano en honor de Karl Von Linneo
(1707-1778). Dicho sistema se basa en dos principios:
- Cada especie de ser vivo (animal, planta, protista o bacteria) recibe un nombre único, en latín, construido con dos palabras (un binomio). Dicho binomio consta de un nombre genérico, que puede ser común a varias especies próximas, y un nombre específico.Por ejemplo: Homo sapiens, Canis familiaris o Pinus pinea
- La agrupación de las especies se hace en estructuras jerárquicas, de mayor o menor categoría. Esto quiere decir que los taxones (unidades de agrupación) de una categoría determinada se agrupan en taxones de superior categoría y comprenden taxones de categoría inferior. En el caso de la especie humana, esto se reflejaría en la siguiente tabla, que recoge las categorías linneanas "clásicas":
| Categoría: | Taxón: | Otros taxones próximos con la misma categoría |
| Reino | Animales (Metazoa) | Plantas, Protistas... |
| Phylum | Cordados | Equinodermos, Hemicordados... |
| Clase | Mamíferos | Arcosaurios, Anfibios... |
| Orden | Primates | Quirópteros, Dermápteros... |
| Familia | Homínidos | Póngidos, Cercopitécidos... |
| Género | Homo | Australopithecus... |
| Especie | Homo sapiens | Homo erectus |
Linneo, por tanto, proporciona un sistema formal de agrupación con una gran capacidad para almacenar información. Pero es preciso preguntarse ahora: ¿Cuál es el criterio del naturalista para agrupar determinados taxones (p.e. especies o familias) dentro de un taxón de categoría superior (p.e. géneros u órdenes)? Cuando a finales del XIX se impuso la idea de evolución,se pudo utilizar con este fin el parentesco filogenético, como luego veremos. Pero, ¿cómo se hacían las agrupaciones antes de este acontecimiento?
El criterio fundamental
también lo proporciona Aristóteles, y se basa en el esencialismo.
El naturalista trata de distinguir, en las características de los
organismos, las que son esenciales (las que definen su "esencia", su "causa
formal") de las que no lo son (caracteres accidentales, no esenciales).
De esta forma una especie determinada, un género o una familia concreta
revelan caracteres "diagnósticos". Estos caracteres diagnósticos,
básicamente morfológicos, permiten agrupar a los individuos
en especies, o a los taxones de inferior categoría en taxones de
categoría superior. La tarea del sistemático sería,
por tanto, la de identificar los caracteres esenciales y realizar agrupaciones
(y clasificaciones) en función de ellos.
Así, decimos de este organismo que es un tiburón (técnicamente: pertenece a los elasmobranquios galeomorfos) porque reúne las características esenciales de los elasmobranquios (endoesqueleto cartilaginoso, dientes de sustitución, escamas placoideas) y las de los galeomorfos (cinco hendiduras branquiales y aleta anal). Por supuesto, cualquier otro ser vivo que reúna los cinco caracteres esenciales antes citados debe ser considerado tiburón (elasmobranquio galeomorfo). Por otro lado, este individuo tiene una serie de caracteres, como la aleta caudal semilunar o el color gris, que no influyen en la consideración de que se trata de un tiburón, aunque pueden ser caracteres esenciales a la hora de determinar el género o la especie.
En este contexto esencialista, de raíz aristotélica, es donde podemos situar el concepto tipológico de especie. En efecto, si decimos que un animal concreto, un individuo, pertenece a la especie Canis familiaris (el perro) es porque reúne una serie de características esenciales para ser considerado como tal (junto con otras no esenciales que hacen de dicho individuo un caniche, un pequinés o un chihuahua).
Este criterio, claro y sencillo, se complicó cuando progresivamente, a lo largo del XIX, se fue imponiendo la idea de evolución. En efecto, si todos los seres vivos son el resultado de un proceso evolutivo, contamos con una nueva variable a la hora de realizar agrupaciones: el parentesco filogenético. En el próximo tema desarrollaremos esta noción. Bástenos ahora señalar que, en efecto, hoy en día existen dos criterios para realizar agrupaciones de taxones:
- La similitud morfológica, es decir, la compartición de características definitorias del taxón. Agrupamos a las aves en el taxón Aves porque tienen plumas, alas, pico córneo, etc..., es decir porque reúnen los caracteres "esenciales" de las aves.
- El parentesco filogenético, es decir, la proximidad de los ancestrales comunes. Agruparemos a las aves en un mismo taxón porque todas comparten un ancestral común relativamente reciente y, desde luego, más reciente que el ancestral que cualquier ave comparte con cualquier otro vertebrado.
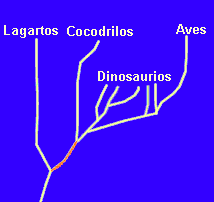 En
el caso anterior no hay conflicto. Las aves estarán agrupadas en
el taxón Aves independientemente del criterio elegido. Pero
en realidad el conflicto entre ambos criterios
puede existir. Un ejemplo muy típico es el de la agrupación
de lagartos, cocodrilos y aves. Si atendemos a criterios morfológicos,
a primera vista lagartos y cocodrilos se parecen más entre sí,
lo que ha justificado su agrupación durante siglos en el taxón
"Reptiles". En cambio, la filogenia del grupo (representada en la
figura) señala que aves y cocodrilos son ramas derivadas de un linaje
común (señalado en naranja) y por tanto, como veremos en
el próximo tema, comparten un mayor grado de parentesco. Dicho parentesco
favorece una agrupación Cocodrilos+Aves en un taxón que se
ha denominado "Arcosaurios". Luego veremos las consecuencias de
este tipo de conflictos.
En
el caso anterior no hay conflicto. Las aves estarán agrupadas en
el taxón Aves independientemente del criterio elegido. Pero
en realidad el conflicto entre ambos criterios
puede existir. Un ejemplo muy típico es el de la agrupación
de lagartos, cocodrilos y aves. Si atendemos a criterios morfológicos,
a primera vista lagartos y cocodrilos se parecen más entre sí,
lo que ha justificado su agrupación durante siglos en el taxón
"Reptiles". En cambio, la filogenia del grupo (representada en la
figura) señala que aves y cocodrilos son ramas derivadas de un linaje
común (señalado en naranja) y por tanto, como veremos en
el próximo tema, comparten un mayor grado de parentesco. Dicho parentesco
favorece una agrupación Cocodrilos+Aves en un taxón que se
ha denominado "Arcosaurios". Luego veremos las consecuencias de
este tipo de conflictos.
La
Nueva Sistemática o Sistemática Evolutiva, una
corriente desarrollada a partir de los años 40 por Ernst Mayr y
Georges Simpson, constató que desde la aceptación de la idea
de la evolución no se habia producido ningún cambio en la
práctica de la Sistemática. Es decir, se seguía trabajando
como en los tiempos de Linneo. La Sistemática Evolutiva propuso
una serie de mejoras metodológicas, tales como:
| La utilización del concepto biológico de especie, basado en propiedades biológicas (la comunidad reproductiva) y no el concepto tipológico, basado en en el esencialismo, como vismos antes. | |
| La utilización de caracteres morfológicos y no morfológicos, y la necesidad de valorar adecuadamente (con métodos estadísticos si es necesario) la variabilidad. | |
| La necesidad de ponderar similitud morfológica y parentesco filogenético en caso de conflicto. |
Estas propuestas fueron bastante criticadas a lo largo de los años 60. Por ejemplo, se señaló que el concepto biológico de especie no era operativo. En la práctica no se comprueba, en la mayoría de los casos, el criterio de interfecundidad. Lo que es peor, especies claramente diferentes, y que no se cruzan en la naturaleza, hibridan sin problemas en condiciones de laboratorio. En realidad, ante las dificultades de la aplicación del concepto biológico de especie, los taxónomos siguen usando criterios tipológicos a la hora de definirlas.
Por otro lado, no quedaba claro cómo había que proceder en caso de conflicto entre similitud y parentesco. También se señaló que los criterios de agrupación en categorías supraespecíficas eran subjetivos. Que una familia determinada comprendiera dos, tres o cuatro géneros no podía justificarse de forma objetiva, y la aceptación de una u otra posibilidad parecía depender de la autoridad del que realizaba la agrupación. De esta forma la práctica sistemática parecía quedar fuera del dominio científico.
Antes esas críticas surgieron dos alternativas que pretendían un método sistemático científico, es decir, objetivo y replicable. La primera de estas alternativas se denominó Feneticismo o Taxonomía Numérica. La segunda, que será la que expondremos a lo largo de este curso, es la Sistemática Filogenética.
Los principios de la Taxonomía Numérica fueron desarrollados por Sokal y Sneath en dos libros publicados en 1963 y 1973. La idea básica era la de resolver el dilema similitud/parentesco en beneficio de la primera. Utilizar sólo la similitud morfológica y cuantificarla en forma de Indices de Similitud o de Distancia. Dichos índices deberían reflejar el parecido global entre los taxones.
No es objetivo de este curso desarrollar los procedimientos feneticistas. Comentaremos que estos procedimientos se basan en el uso de matrices en las que se incluyen valores numéricos de determinados atributos de las unidades a agrupar, unidades llamadas OTUs (de operative taxonomical unity) que pueden ser individuos, muestras de poblaciones o especies, etc. Los atributos pueden ser medidas, proporciones, recuentos o caracteres semicuantitativos (p.e. 0=ausencia, 1=presencia). La idea es que cuanto más similares sean dos OTUs, menor será la diferencia global entre los valores de sus atributos. De esta forma se pueden definir índices de distancia fenética, como por ejemplo la media cuadrática de las diferencias entre cada par de atributos. En el caso de identidad total entre dos OTUs dicho índice valdría 0, y será mayor cuanto más disimilares sean los OTUs.
Ejemplo de matriz
de datos:
|
|
OTU1 |
OTU2 |
OTU3 |
|
Car. 1 |
24 |
26 |
29 |
|
Car. 2 |
12,2 |
14 |
15 |
|
Car. 3 |
2 |
3 |
3 |
Ejemplo de Indices de distancia:
|
|
OTU1 |
OTU2 |
|
OTU2 |
1,89 |
|
|
OTU3 |
3,34 |
1,49 |
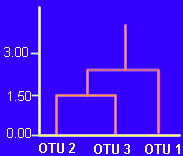
Los OTUs 2 y 3,
con mayor parecido global, se sitúan a una distancia de 1,49. La
distancia del OTU 1 al grupo (OTU2+OTU3) es igual a la media aritmética
de las distancias a cada uno de sus componentes, es decir: (1,89+3,34)/2
= 2,61.
La situación en la que se enmarca el nacimiento de la Sistemática Filogenética es, por tanto, una sistemática "evolutiva" que a pesar de su nombre seguía utilizando procedimientos básicamente esencialistas o preevolutivos. El concepto biológico de especie resultaba difícil de aplicar en la práctica, por lo que se aplicaba el concepto tipológico clásico. Se planteaban conflictos irresolubles entre similitud y filogenia. Y en última instancia la clasificación contaba con un componente fuertemente subjetivo. La alternativa feneticista, por otra parte, abandonaba no ya el concepto de evolución, sino cualquier sentido biológico en la clasificación. La similitud se conviertía en la única propiedad relevante, con lo que trabajar con seres vivos no resultaba diferente a trabajar con cualquier tipo de objetos. Veremos en los próximos temas como la Sistemática Filogenética intenta resolver estos problemas.
Pero antes podemos pasar a realizar algunos ejercicios:
